Symbiotic associations between endophytic fungi and Chenopodium quinoa have been reported to be beneficial for increasing tolerance to drought and soil salinity, being this the only crop grown in the Southern Altiplano of Bolivia. The symbiotic interaction of endophytic fungi was investigated, considering their detrimental and mutualistic effects. 38 strains of endophytic fungi from the fungal collection of the Faculty of Agronomy were used. The percentage of germination and root length were measured in vitro (4 days). The percentage of emergence (7 days) and plant height (14 days) in pots were also evaluated. The fungi that did not impair germination were strains VP42, VP44, Alternaria sp. VP37, Fusarium sp. VP05 and Fusarium sp. VP30. Fungi that stimulated a significant increase in radicle longitudinal growth were Fusarium sp. VP35, Alternaria sp. VP37 and strain VP18. Fungi that caused a high mortality rate during the emergence phase were strain VP01, Alternaria sp. VP15, Fusarium sp. VP02, Fusarium sp. VP07, Fusarium sp. VP08, Fusarium sp. VP12, Fusarium sp. VP23 and Fusarium sp. VP36. The endophytic strains Alternaria sp. VP37, Fusarium sp. VP35 and strain VP18 stimulated superior seedling growth. The present research work reveals that some endophytic fungi of the Alternaria and Fusarium genera can behave as pathogens during the germination stage, while others have the function of promoting quinoa growth.
| Published in | American Journal of BioScience (Volume 13, Issue 1) |
| DOI | 10.11648/j.ajbio.20251301.11 |
| Page(s) | 1-10 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2025. Published by Science Publishing Group |
Alternaria, Fusarium, Endophytic Fungus, Pathogenesis, Quinoa
Specimens | Strain code | Host | Place |
|---|---|---|---|
Alternaria sp. | VP15 | Quinoa root | Southern Altiplano - Bolivia |
Alternaria sp. | VP24 | Quinoa root | Southern Altiplano - Bolivia |
Alternaria sp. | VP37 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP12 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP11 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP16 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP02 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP21 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP22 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP23 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP03 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP30 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP35 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP36 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP39 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP40 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP41 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP43 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP05 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP06 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP07 | Quinoa root | Southern Altiplano - Bolivia |
Fusarium sp. | VP08 | Quinoa root | Southern Altiplano - Bolivia |
Penicillium sp. | VP32 | Quinoa root | Southern Altiplano - Bolivia |
Penicillium sp. | VP38 | Quinoa root | Southern Altiplano - Bolivia |
Unknown | VP01 | Quinoa root | Southern Altiplano - Bolivia |
Unknown | VP14 | Quinoa root | Southern Altiplano - Bolivia |
Unknown | VP18 | Quinoa root | Southern Altiplano - Bolivia |
Unknown | VP19 | Quinoa root | Southern Altiplano - Bolivia |
Unknown | VP20 | Quinoa root | Southern Altiplano - Bolivia |
Unknown | VP28 | Quinoa root | Southern Altiplano - Bolivia |
Unknown | VP29 | Quinoa root | Southern Altiplano - Bolivia |
Unknown | VP31 | Quinoa root | Southern Altiplano - Bolivia |
Unknown | VP33 | Quinoa root | Southern Altiplano - Bolivia |
Unknown | VP34 | Quinoa root | Southern Altiplano - Bolivia |
Unknown | VP04 | Quinoa root | Southern Altiplano - Bolivia |
Unknown | VP42 | Quinoa root | Southern Altiplano - Bolivia |
Unknown | VP44 | Quinoa root | Southern Altiplano - Bolivia |
Unknown | VP09 | Quinoa root | Southern Altiplano - Bolivia |
EF | Endophytic Fungi |
MS | Murashige and Skoog |
PDA | Potato Dextrose Agar |
IAA | Indole Acetic Acid |
GA | Gibberellic Acid |
| [1] | A. Vega-Gálvez, M. Miranda, J. Vergara, E. Uribe, L. Puente, y E. A. Martínez, «Nutrition facts and functional potential of quinoa (Chenopodium quinoa willd.), an ancient Andean grain: a review», J. Sci. Food Agric., vol. 90, n.o 15, pp. 2541-2547, dic. 2010, |
| [2] | D. Bazile, «Global Trends in the Worldwide Expansion of Quinoa Cultivation», Biol. Life Sci. Forum, vol. 15, n.o 1, p. 13, 2023, [En línea]. Disponible en: |
| [3] | D. Bazile, S.-E. Jacobsen, y A. Verniau, «The Global Expansion of Quinoa: Trends and Limits», Front. Plant Sci., vol. 7, p. 622, 2016, |
| [4] | M. Walsh-Dilley, «Resilience compromised: Producing vulnerability to climate and market among quinoa producers in Southwestern Bolivia», Glob. Environ. Chang., vol. 65, p. 102165, 2020, |
| [5] | F. Lutzoni et al., «Contemporaneous radiations of fungi and plants linked to symbiosis», Nat. Commun., vol. 9, n.o 1, p. 5451, 2018, |
| [6] | C. Strullu-Derrien, M. A. Selosse, P. Kenrick, y F. M. Martin, «The origin and evolution of mycorrhizal symbioses: from palaeomycology to phylogenomics», New Phytol., vol. 220, n.o 4, pp. 1012-1030, 2018, |
| [7] | N. Vigneron, G. V. Radhakrishnan, y P. M. Delaux, «What have we learnt from studying the evolution of the arbuscular mycorrhizal symbiosis?», Curr. Opin. Plant Biol., vol. 44, pp. 49-56, 2018, |
| [8] | M. A. Hassani, P. Durán, y S. Hacquard, «Microbial interactions within the plant holobiont», Microbiome, pp. 6-58, 2018, |
| [9] | J. Mercado-Blanco, «Life of Microbes Inside the Plant», en Principles of Plant-Microbe Interactions: Microbes for Sustainable Agriculture, B. Lugtenberg, Ed. Springer, 2015, pp. 25-32. |
| [10] | W. K. Mousa y M. N. Raizada, «The diversity of anti-microbial secondary metabolites produced by fungal endophytes: an interdisciplinary perspective», Front. Microbiol., vol. 4, p. 65, 2013, |
| [11] | Z.-L. Yuan, C.-L. Zhang, y F.-C. Lin, «Role of Diverse Non-Systemic Fungal Endophytes in Plant Performance and Response to Stress: Progress and Approaches», J. Plant Growth Regul., vol. 29, n.o 1, pp. 116-126, 2010, |
| [12] | P. Bonfante y A. Genre, «Mechanisms underlying beneficial plant - Fungus interactions in mycorrhizal symbiosis», Nat. Commun., p. 1: 48, 2010, |
| [13] | F. A. Decunta, L. I. Pérez, D. P. Malinowski, M. A. Molina-Montenegro, y P. E. Gundel, «A Systematic Review on the Effects of Epichloë Fungal Endophytes on Drought Tolerance in Cool-Season Grasses», Front. Plant Sci., vol. 12, p. 644731, 2021, |
| [14] | K. M. Thambugala, D. A. Daranagama, A. J. L. Phillips, S. D. Kannangara, y I. Promputtha, «Fungi vs. Fungi in Biocontrol: An Overview of Fungal Antagonists Applied Against Fungal Plant Pathogens», Front. Cell. Infect. Microbiol., vol. 10, n.o November, p. 604923, 2020, |
| [15] | P. Trivedi, J. E. Leach, S. G. Tringe, T. Sa, y B. K. Singh, «Plant-microbiome interactions: from community assembly to plant health», Nat. Rev. Microbiol., vol. 18, n.o 11, pp. 607-621, 2020, |
| [16] | A. E. Fadiji y O. O. Babalola, «Exploring the potentialities of beneficial endophytes for improved plant growth», Saudi J. Biol. Sci., vol. 27, n.o 12, pp. 3622-3633, 2020, |
| [17] | P. R. Hardoim et al., «The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes», Microbiol. Mol. Biol. Rev., vol. 79, n.o 3, pp. 293-320, 2015, |
| [18] | K. Selim, «Biology of Endophytic Fungi», Curr. Res. Environ. Appl. Mycol., vol. 2, n.o 1, pp. 31-82, 2012, |
| [19] | V. V. Kumar, «Plant Growth-Promoting Microorganisms: Interaction with Plants and Soil», en Plant, Soil and Microbes, vol. 1, K. R. Hakeem, M. S. Akhtar, y S. N. A. Abdullah, Eds. Springer International Publishing Switzerland, 2016, pp. 1-16. |
| [20] | M. González-Teuber, R. A. Contreras, G. E. Zúñiga, D. Barrera, y L. Bascuñán-Godoy, «Synergistic Association With Root Endophytic Fungi Improves Morpho-Physiological and Biochemical Responses of Chenopodium quinoa to Salt Stress», Front. Ecol. Evol., vol. 9, p. 787318, 2022, |
| [21] | M. González-Teuber, C. Vilo, y L. Bascuñán-Godoy, «Molecular characterization of endophytic fungi associated with the roots of Chenopodium quinoa inhabiting the Atacama Desert, Chile», Genomics Data, vol. 11, pp. 109-112, 2017, |
| [22] | G. Berg, P. Kusstatscher, A. Abdelfattah, T. Cernava, y K. Smalla, «Microbiome Modulation-Toward a Better Understanding of Plant Microbiome Response to Microbial Inoculants», Front. Microbiol., vol. 12, p. 650610, 2021, |
| [23] | Y. A. Millet et al., «Innate immune responses activated in Arabidopsis roots by microbe-associated molecular patterns», Plant Cell, vol. 22, n.o 3, pp. 973-990, 2010, |
| [24] | A. C. Perez, J. S. Rojas, y H. M. Vale, «Biologia y perspectiva de microorganismos endófitos asociados a plantas», Rev. Colomb. Cienc. Anim. - RECIA, vol. 1, n.o 2, pp. 286-301, 2009, |
| [25] | A. Zuccaro et al., «Endophytic life strategies decoded by genome and transcriptome analyses of the mutualistic root symbiont Piriformospora indica», PLoS Pathog., vol. 7, n.o 10, p. e1002290, 2011, |
| [26] | I. Khalmuratova, D. H. Choi, H. J. Yoon, T. M. Yoon, y J. G. Kim, «Diversity and plant growth promotion of fungal endophytes in five halophytes from the buan salt marsh», J. Microbiol. Biotechnol., vol. 31, n.o 3, pp. 408-418, 2021, |
| [27] | A. E. Arnold y F. Lutzoni, «Diversity and host range of foliar fungal endophytes: are tropical leaves biodiversity hotspots?», Ecology, vol. 88, n.o 3, pp. 541-549, 2007, |
| [28] | S. Kusari, S. P. Pandey, y M. Spiteller, «Untapped mutualistic paradigms linking host plant and endophytic fungal production of similar bioactive secondary metabolites», Phytochemistry, vol. 91, pp. 81-87, 2012, |
| [29] | N. M. Chauhan, A. D. Gutama, y A. Aysa, «Endophytic fungal diversity isolated from different agro-ecosystem of Enset (Ensete ventericosum) in Gedeo zone, SNNPRS, Ethiopia», BMC Microbiol., vol. 19, n.o 1, p. 172, 2019, [En línea]. Disponible en: |
| [30] | J. G. Harrison y E. A. Griffin, «The diversity and distribution of endophytes across biomes, plant phylogeny and host tissues: how far have we come and where do we go from here?», Environ. Microbiol., vol. 22, n.o 6, pp. 2107-2123, 2020, |
| [31] | L. Jin et al., «Diversity and Biocontrol Potential of Culturable Endophytic Fungi in Cotton», Front. Microbiol., vol. 12, p. 698930, 2021, |
| [32] | K. L. Rana et al., «Endophytic microbes: biodiversity, plant growth-promoting mechanisms and potential applications for agricultural sustainability», Antonie Van Leeuwenhoek, vol. 113, n.o 8, pp. 1075-1107, 2020, |
| [33] | R. E. Sánchez-Fernández et al., «Hongos endófitos: fuente potencial de metabolitos secundarios bioactivos con utilidad en agricultura y medicina», Tip Rev. Espec. en Ciencias Químico-Biológicas, vol. 16, n.o 2, pp. 132-146, 2013, |
| [34] | J. A. Castillo, G. Conde, M. Claros, y N. Ortuño, «Diversity of cultivable microorganisms associated with Quinoa (Chenopodium quinoa) and their potential for plant growth-promotion», Bionatura, vol. 7, n.o 2, p. 61, 2022, |
| [35] | J. O. de Carvalho, V. Broll, A. H. S. Martinelli, y F. C. Lopes, «Endophytic fungi: positive association with plants», en Molecular Aspects of Plant Beneficial Microbes in Agriculture, V. Sharma, R. Salwan, y L. K. Tawfeeq Al-Ani, Eds. India: Academic Press, 2020, pp. 321-332. [En línea]. Disponible en: |
| [36] | N. Ortuño, J. A. Castillo, y M. Claros, «Symbiotic Native Microorganisms of Quinoa in the Bolivian Altiplano», en Biology and Biotechnology of Quinoa, A. Varma, Ed. Springer Nature Singapore, 2021, pp. 131-151. [En línea]. Disponible en: |
| [37] | P. W. Crous et al., «Fusarium: more than a node or a foot-shaped basal cell», Stud. Mycol., vol. 98, p. 100116, 2021, |
| [38] | M. R. Panuccio, S. E. Jacobsen, S. S. Akhtar, y A. Muscolo, «Effect of saline water on seed germination and early seedling growth of the halophyte quinoa», AoB Plants, vol. 6, p. plu047, 2014, |
| [39] | A. Naureen, F. ul H. Nasim, M. S. Choudhary, M. Ashraf, F. M. W. Grundler, y A. S. S. Schleker, «A new endophytic fungus CJAN1179 isolated from the Cholistan desert promotes lateral root growth in Arabidopsis and produces IAA through tryptophan-dependent pathway», Arch. Microbiol., vol. 204, n.o 3, p. 181, 2022, |
| [40] | N. C. Baron, A. de Souza Pollo, y E. C. Rigobelo, «Purpureocillium lilacinum and Metarhizium marquandii as plant growth-promoting fungi», PeerJ, n.o 8, p. e9005, 2020, |
| [41] | G. Berg, «Plant-microbe interactions promoting plant growth and health: Perspectives for controlled use of microorganisms in agriculture», Appl. Microbiol. Biotechnol., vol. 84, n.o 1, pp. 11-18, 2009, |
| [42] | L. R. Jaber y J. Enkerli, «Fungal entomopathogens as endophytes: can they promote plant growth?», Biocontrol Sci. Technol., vol. 27, n.o 1, pp. 28-41, 2017, |
| [43] | A. L. Khan, J. Hussain, A. Al-Harrasi, A. Al-Rawahi, y I.-J. Lee, «Endophytic fungi: resource for gibberellins and crop abiotic stress resistance», Crit. Rev. Biotechnol., vol. 35, pp. 62-74, 2015. |
| [44] | N. C. Baron y E. C. Rigobelo, «Endophytic fungi: a tool for plant growth promotion and sustainable agriculture», Mycology, vol. 13, n.o 1, pp. 39-55, 2022, |
| [45] | L. Yan, J. Zhu, X. Zhao, J. Shi, C. Jiang, y D. Shao, «Beneficial effects of endophytic fungi colonization on plants», Appl. Microbiol. Biotechnol., vol. 103, n.o 8, pp. 3327-3340, 2019, |
| [46] | M. González-Teuber, A. Urzuá, P. Plaza, y L. Bascuñán-Godoy, «Effects of root endophytic fungi on response of Chenopodium quinoa to drought stress», Plant Ecol., vol. 219, pp. 231-240, 2018, |
| [47] | L. Bilal et al., «Plant growth promoting endophytic fungi Asprgillus fumigatus TS1 and Fusarium proliferatum BRL1 produce gibberellins and regulates plant endogenous hormones», Symbiosis, vol. 76, pp. 117-127, 2018, [En línea]. Disponible en: |
| [48] | S. Roy, A. Sarma, S. Paul, D. K. Jha, y K. Tayung, «Plant growth-promoting traits and activation of defense enzymes intraditional rice variety by fungal endophytes isolated from seeds ofindigenous rice cultivars of Northeast India», South African J. Bot., vol. 160, pp. 483-492, 2023, [En línea]. Disponible en: |
| [49] | N. A. Elhamouly et al., «The hidden power of secondary metabolites in plant-fungi interactions and sustainable phytoremediation», Front. Plant Sci., vol. 13, p. 1044896, 2022, |
| [50] | P. Tiwari y H. Bae, «Endophytic Fungi: Key Insights, Emerging Prospects, and Challenges in Natural Product Drug Discovery», Microorganisms, vol. 10, p. 360, 2022, |
| [51] | S. E. Smith, E. Facelli, S. Pope, y F. A. Smith, «Plant performance in stressful environments: interpreting new and established knowledge of the roles of arbuscular mycorrhizas», Plant Soil, vol. 326, pp. 3-20, 2010, |
| [52] | H. A. Bruns, «Controlling Aflatoxin and Fumonisin in Maize by Crop Management», J. Toxicol., vol. 22, n.o 2-3, pp. 153-173, 2003, |
| [53] | M. Kottb, T. Gigolashvili, D. K. Großkinsky, y B. Piechulla, «Trichoderma volatiles effecting Arabidopsis: from inhibition to protection against phytopathogenic fungi», Front. Microbiol., vol. 6, p. 995, 2015, |
| [54] | A. M. Abdelaziz, D. A. El-Wakil, M. S. Attia, O. M. Ali, H. AbdElgawad, y A. H. Hashem, «Inhibition of Aspergillus flavus Growth and Aflatoxin Production in Zea mays L. Using Endophytic Aspergillus fumigatus», J. Fungi, vol. 8, n.o 5, p. 482, 2022, |
| [55] | J. P. Louw y L. Korsten, «Pathogenic Penicillium spp. on apple and pear», Plant Dis., vol. 98, n.o 5, pp. 590-598, 2014, |
APA Style
Pérez, V. P., Fabiana, C. M. C., Quisbert, F. P. (2025). Evaluation of Growth Promotion and Pathogenicity of Endophytic Fungi from the Root of Chenopodium Quinoa Willd. American Journal of BioScience, 13(1), 1-10. https://doi.org/10.11648/j.ajbio.20251301.11
ACS Style
Pérez, V. P.; Fabiana, C. M. C.; Quisbert, F. P. Evaluation of Growth Promotion and Pathogenicity of Endophytic Fungi from the Root of Chenopodium Quinoa Willd. Am. J. BioScience 2025, 13(1), 1-10. doi: 10.11648/j.ajbio.20251301.11
@article{10.11648/j.ajbio.20251301.11,
author = {Victor Paco Pérez and Crespo Melgar Carla Fabiana and Fernando Pacasa Quisbert},
title = {Evaluation of Growth Promotion and Pathogenicity of Endophytic Fungi from the Root of Chenopodium Quinoa Willd
},
journal = {American Journal of BioScience},
volume = {13},
number = {1},
pages = {1-10},
doi = {10.11648/j.ajbio.20251301.11},
url = {https://doi.org/10.11648/j.ajbio.20251301.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ajbio.20251301.11},
abstract = {Symbiotic associations between endophytic fungi and Chenopodium quinoa have been reported to be beneficial for increasing tolerance to drought and soil salinity, being this the only crop grown in the Southern Altiplano of Bolivia. The symbiotic interaction of endophytic fungi was investigated, considering their detrimental and mutualistic effects. 38 strains of endophytic fungi from the fungal collection of the Faculty of Agronomy were used. The percentage of germination and root length were measured in vitro (4 days). The percentage of emergence (7 days) and plant height (14 days) in pots were also evaluated. The fungi that did not impair germination were strains VP42, VP44, Alternaria sp. VP37, Fusarium sp. VP05 and Fusarium sp. VP30. Fungi that stimulated a significant increase in radicle longitudinal growth were Fusarium sp. VP35, Alternaria sp. VP37 and strain VP18. Fungi that caused a high mortality rate during the emergence phase were strain VP01, Alternaria sp. VP15, Fusarium sp. VP02, Fusarium sp. VP07, Fusarium sp. VP08, Fusarium sp. VP12, Fusarium sp. VP23 and Fusarium sp. VP36. The endophytic strains Alternaria sp. VP37, Fusarium sp. VP35 and strain VP18 stimulated superior seedling growth. The present research work reveals that some endophytic fungi of the Alternaria and Fusarium genera can behave as pathogens during the germination stage, while others have the function of promoting quinoa growth.
},
year = {2025}
}
TY - JOUR T1 - Evaluation of Growth Promotion and Pathogenicity of Endophytic Fungi from the Root of Chenopodium Quinoa Willd AU - Victor Paco Pérez AU - Crespo Melgar Carla Fabiana AU - Fernando Pacasa Quisbert Y1 - 2025/01/09 PY - 2025 N1 - https://doi.org/10.11648/j.ajbio.20251301.11 DO - 10.11648/j.ajbio.20251301.11 T2 - American Journal of BioScience JF - American Journal of BioScience JO - American Journal of BioScience SP - 1 EP - 10 PB - Science Publishing Group SN - 2330-0167 UR - https://doi.org/10.11648/j.ajbio.20251301.11 AB - Symbiotic associations between endophytic fungi and Chenopodium quinoa have been reported to be beneficial for increasing tolerance to drought and soil salinity, being this the only crop grown in the Southern Altiplano of Bolivia. The symbiotic interaction of endophytic fungi was investigated, considering their detrimental and mutualistic effects. 38 strains of endophytic fungi from the fungal collection of the Faculty of Agronomy were used. The percentage of germination and root length were measured in vitro (4 days). The percentage of emergence (7 days) and plant height (14 days) in pots were also evaluated. The fungi that did not impair germination were strains VP42, VP44, Alternaria sp. VP37, Fusarium sp. VP05 and Fusarium sp. VP30. Fungi that stimulated a significant increase in radicle longitudinal growth were Fusarium sp. VP35, Alternaria sp. VP37 and strain VP18. Fungi that caused a high mortality rate during the emergence phase were strain VP01, Alternaria sp. VP15, Fusarium sp. VP02, Fusarium sp. VP07, Fusarium sp. VP08, Fusarium sp. VP12, Fusarium sp. VP23 and Fusarium sp. VP36. The endophytic strains Alternaria sp. VP37, Fusarium sp. VP35 and strain VP18 stimulated superior seedling growth. The present research work reveals that some endophytic fungi of the Alternaria and Fusarium genera can behave as pathogens during the germination stage, while others have the function of promoting quinoa growth. VL - 13 IS - 1 ER -
Technical University of Oruro, Faculty of Agricultural and Natural Sciences, Oruro, Plurinational State of Bolivia
Mayor University of San Andrés, Faculty of Pharmaceutical and Biological Sciences, La Paz, Plurinational State of Bolivia
Technical University of Oruro, Faculty of Agricultural and Natural Sciences, Oruro, Plurinational State of Bolivia
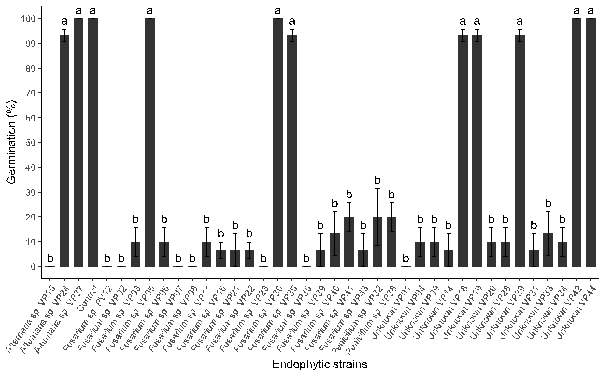
Figure 1. Quinoa germination can be influenced by inoculation with endophytic fungi.
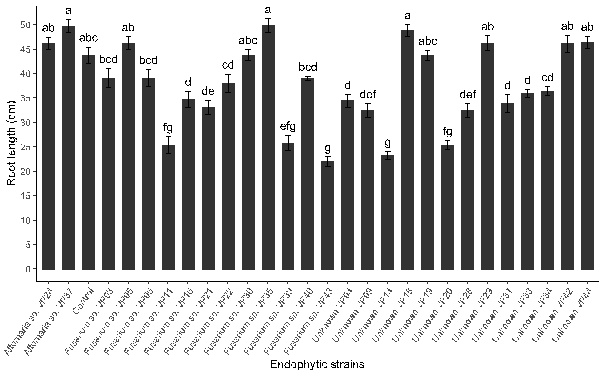
Figure 2. Root development of quinoa can be influenced by inoculation with endophytic fungi.
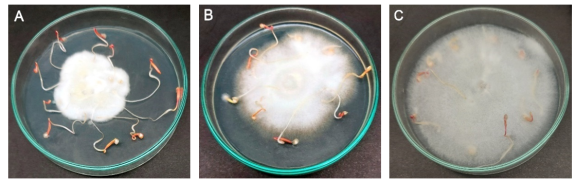
Figure 3. In vitro test: Endophytic fungi effect on quinoa root length. (A) Fungus observed to be beneficial as it promotes root growth, (B) Fungi has a moderately detrimental effect on root development, and (C) certain endophytic fungi are detrimental to root development.
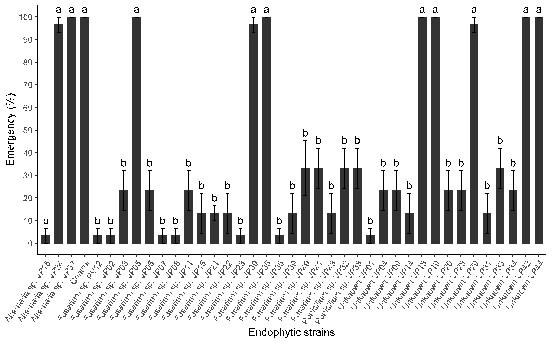
Figure 4. Effect of inoculation with endophytic fungi in the emergence phase of quinoa.
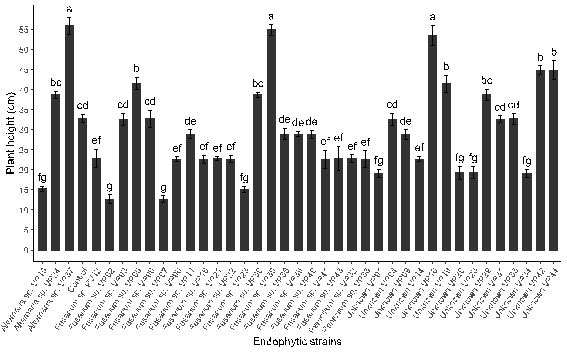
Figure 5. The effect of inoculation with endophytic fungi on the height growth of quinoa plants.

Figure 6. Impact of endophytic fungi inoculation on quinoa growth. In (A), a positive boost in development is evident due to the mutualistic strain. On the other hand, in (B), it is observed that the endophytic fungus has a moderate detrimental effect, while in (C), it is shown that this strain is highly detrimental due to its pathogenic behavior.

